





Il bilancio negativo dei donatori di gruppi metilici (NMDB)
Nella prima parte di questo articolo abbiamo descritto il generico bilancio proteico negativo (NPB) della bovina da latte. Gli amminoacidi sono 20 e sono suddivisi più o meno al 50% tra essenziali (EAA), ossia che la bovina non può sintetizzare in quantità sufficiente ma che devono essere messi a disposizione dalle riserve corporee o assunti tramite la dieta, e non essenziali (NEAA) cioè che possono essere sintetizzati nell’organismo con poco o nessun rischio di carenze. Questa ripartizione non è poi così rigida ma, per necessità di trattazione, possiamo classificare come EAA: lisina, metionina, valina, arginina, isoleucina, istidina, triptofano, leucina, treonina e fenilalanina. Tra i NEAA troviamo invece: glicina, serina, acido glutammico, cisteina, tirosina, prolina, alanina, acido aspartico, glutammina e idrossiprolina. Oltre che in questi due gruppi di base, gli amminoacidi possono anche essere suddivisi in glicogenetici (partecipano alla sintesi del glucosio), insulinogenetici e chetogenetici. Con il progredire delle ricerche sul metabolismo della bovina da latte si sta assegnando via via un ruolo sempre più chiaro nel metabolismo ai singoli amminoacidi e alle loro interazioni.
Di certo si sa che:
- L’uptake di EAA della mammella è in eccesso per isoleucina, leucina, lisina e valina.
- La quantità di NEAA necessari per la sintesi della proteina del latte apportati con la dieta non è sufficiente per tutte le tipologie di aminoacidi.
- La mammella sintetizza NEAA dagli EAA captati in eccesso.
- L’uptake di lisina per sintetizzare NEAA ne può causare la carenza.
- L’estrazione di prolina, acido glutammico e acido aspartico è bassa se comparata con la quantità richiesta dalla mammella.
- NEAA come la prolina e l’acido glutammico sono limitanti per la sintesi di proteine nel latte anche se sono sintetizzati nella mammella.
- La quantità di arginina estratta dalla mammella è 2-4 volte maggiore di quella secreta nel latte.
- L’ornitina e la citrullina vengono estratte, ma non le troviamo nella proteina del latte.
- Arginina e ornitina sono precursori nella mammella di prolina e acido glutammico.
- La quantità di fenilalanina e metionina che troviamo nel latte è maggiore del 20-30% rispetto a quella estratta dalla mammella.
- Fenilalanina, metionina, lisina, istidina e treonina sono i cinque aminoacidi essenziali utilizzati per il completamento della ghiandola mammaria.
- Arginina, valina, leucina e isoleucina vengono estratti dalla mammella in quantità superiore all’output nella proteina del latte.
- Prolina, glutammato e aspartato non sono estratti dal sangue in quantità sufficiente per le esigenze della mammella.
Molto si conosce oggi sul ruolo della metionina. Questo amminoacido, classificato tra gli EAA, rappresenta nel latte bovino circa il 5.5% degli EAA totali e dal 2.48 al 3.32% della caseina. La metionina di cui la bovina da latte può disporre è apportata dalla proteina metabolizzabile (MP), costituita in parte da quella microbica e in parte da quella che deriva dalla quota indegradata delle proteine ingerite nella dieta. Un’altra fonte di metionina deriva dalle proteine labili, o meglio quelle muscolari. Il 50% della metionina utilizzata dalla bovina da latte è sintetizzata a partire dalla rimetilazione dell’omocisteina. La metionina, tramite l’enzima metionina adenosil-tranferasi, reagisce con una molecola di ATP trasformandosi in S-adenosil metionina (SAM). Questa molecola è piuttosto reattiva e tende a donare ad un’altra molecola il gruppo metilico CH3+ che si trova legato allo zolfo (transmetilazione). Questa reazione è catalizzata dall’enzima metiltransferasi.
Di questa “donazione” di gruppi metilici possono usufruire DNA, RNA, lipidi, proteine e circa 50 reazioni metaboliche molto importanti.
Più del 30% della metionina assorbita viene utilizzata per la sintesi della colina. La metionina ha un ruolo diretto nella sintesi dell’apolipoproteina B e della fosfatidilcolina. La disponibilità nella dieta di folati, vitamina B12, colina e betaina influenza i fabbisogni di metionina. Nella figura sottostante viene rappresentato il pathway metabolico dei gruppi metilici e l’interazione tra alcune molecole (cerchiate di verde) che possono essere aggiunte come additivi, necessariamente rumino-protetti, nelle diete delle bovine in lattazione ma anche nelle linee high genetic merit (HGM) delle pecore, delle capre e delle bufale da latte.
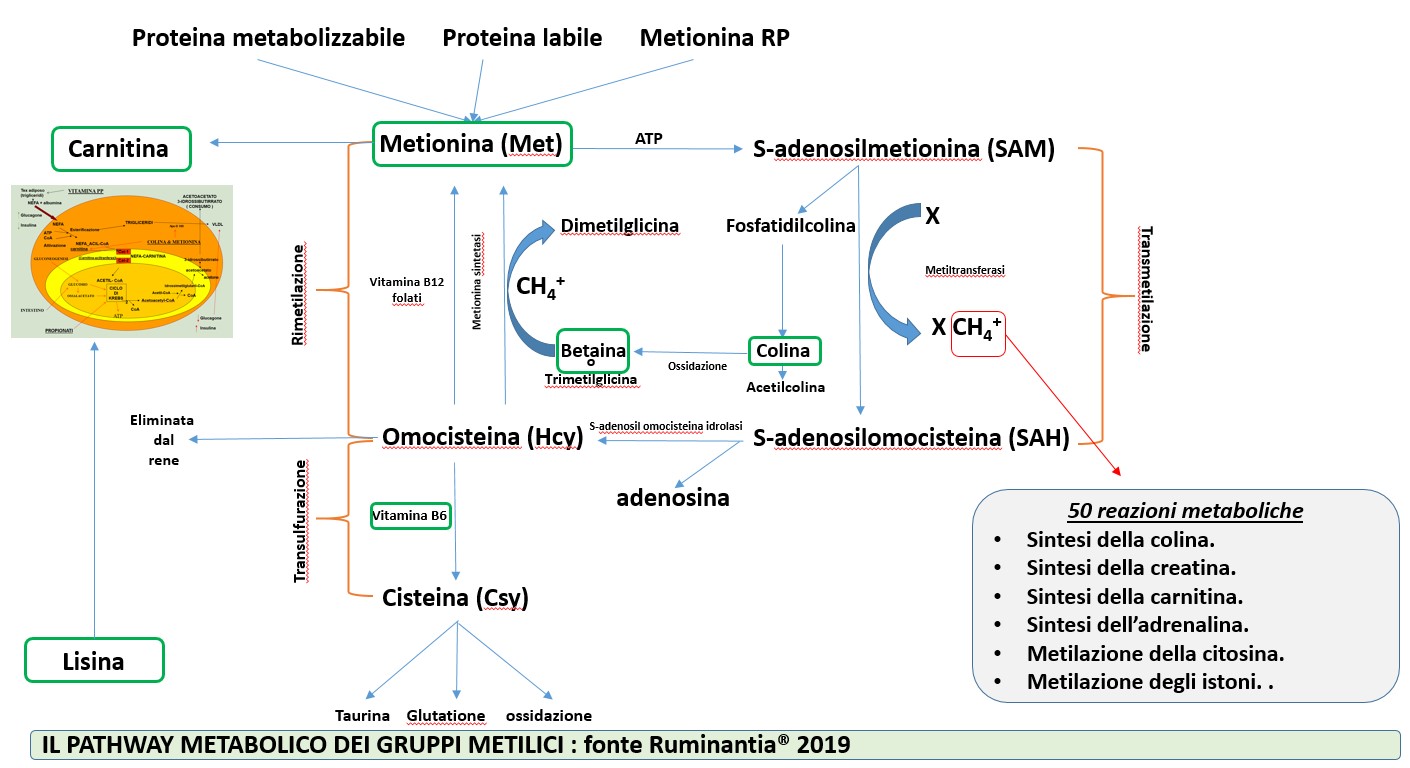
Molto interessante è il ruolo che i gruppi metilici hanno nell’espressione genetica. Sappiamo che la selezione naturale o artificiale plasma gli esseri viventi in una continua e inesorabile ricerca del miglior adattamento possibile all’ambiente circostante. Quest’ultimo però può modulare l’espressione del patrimonio genetico degli esseri viventi attraverso l’inibizione o meno dell’espressione di alcuni geni. Gli adulti (maschi e femmine) possono trasmettere tutto ciò alle generazioni successive. I gruppi metilici hanno un ruolo essenziale in questo processo: la metilazione della citosina provoca il “silenziamento” di alcuni geni, mentre l’acetilazione degli istoni rende accessibile parte del DNA alla trascrizione dell’RNA. Questo meccanismo, chiamato epigenetica, “descrive” letteralmente ciò che avviene in organismi o cellule identiche che hanno una diversa espressione fenotipica, senza un cambiamento della sequenza genetica. Pertanto l’ambiente, attraverso la metilazione e l’acetilazione, può determinare un imprinting sugli animali che viene trasmesso alle generazioni successive in modo da renderle ancora più adatte alle peculiarità dell’ambiente in cui vivono.
Conclusioni
Gli animali d’allevamento stanno subendo ormai da millenni la pressione selettiva dell’uomo per renderli sempre più rispondenti alle proprie esigenze e all’ambiente dove ha deciso di allevarli. La nutrizione e le sue alterazioni come il NEBAL, NPB e NMDB e, più in generale, le malattie metaboliche nel periodo del ciclo produttivo della bovina e dei ruminanti da latte, occorrono principalmente al momento della riproduzione. Questa coincidenza temporale è in grado d’imprimere modificazioni profonde, o meglio epigenetiche, al patrimonio genetico di questi animali. I tanti difetti enzimatici che inevitabilmente si stanno accumulando negli animali d’allevamento e le modificazione genetiche indotte da turbe metaboliche hanno un sicuro impatto negativo sulla loro longevità funzionale e sulla produttività. Il ricorso a farmaci come ormoni ed antibiotici deve essere necessariamente limitato all’indispensabile mentre i farmaci “metabolici” e la nutrizione clinica e funzionale hanno, e avranno, un ruolo sempre più importante.
Rubrica a cura di Vetagro
Contatta Vetagro: info@vetagro.com
Visita il sito: www.vetagro.com




