





Caratterizzazione fenotipica della produzione di latte e dei tratti qualitativi in un’ampia popolazione di bufale.
Sono stati raccolti 1.414.449 record test-day (TD) da 106.388 bufali allevati in 386 aziende dell’Italia centrale e meridionale. Per ciascun record erano disponibili, oltre alle informazioni sulla quantità di latte (Milk Yield – MY), data di nascita e di parto, giorni in lattazione e ordine di parto, e anche le informazioni relative alle caratteristiche qualitative del latte, quali il contenuto in grasso (Fat Content – FC), in proteine (Protein content – PC), in lattosio (Lactose content – LC) e in cellule somatiche (SCC content). Per le bufale munte una sola volta al giorno, la stima del MY è stata ottenuta mediante specifici coefficienti. Le caratteristiche qualitative del latte (FC, PC, LC) sono state ottenute su ciascun campione utilizzando modelli di predizione specifici per bufali implementati nel MilkoScan™ FT6000 (Foss Electric A/S, Hillerød, Danimarca) del laboratorio del latte ARAC di Benevento. Il conteggio delle cellule somatiche (SCC, cellule/mL) è stato determinato tramite Fossomatic™ (Foss Electric A/S, Hillerød, Danimarca), mentre per il latte corretto standardizzato a 740 kcal (ECM, kg/d) è stata utilizzata la seguente formula:
ECM = MY × {[(FC × MY − 40) + (PC × MY − 31)] × 0.01155 + 1}
Al fine di effettuare una corretta analisi dei dati, è stato effettuato un editing del database raccolto. In particolare, le analisi sui controlli quali-quantitativi di latte per MY, FC, PC, LC, and ECM sono state effettuate escludendo gli outliers (media ± 3 SD), mentre i valori di SCC che hanno presentato variazioni da 1000 a 999.000 cellule/ml, sono stati convertiti in somatic cell score (SCS), al fine di ottenere una normalizzazione della distribuzione dei dati (curva di Gauss), in base alla seguente formula:
SCS = 3 + log2(SCC/100,000)
Sono stati presi in cosiderazione soltanto i TD compresi nel range di giorni in lattazione (Days in milk – DIM) tra 5 e 400 e solo le bufale fino al 6° parto. Tale parametro è stato scelto in quanto rappresenta la vita media produttiva degli animali negli allevamenti bufalini in Italia. Un ulteriore editing è stato poi effettuato all’interno di questi dati, con lo scopo di escludere bufale che presentassero un’età al di fuori dei seguenti range per ciascun ordine di parto: 3,32 ± 1,28, 4,66 ± 1,29, 5,91 ± 1,35, 7,08 ± 1,39, 8,24 ± 1,48, and 9,34 ± 1,52 anni per gli animali di primo, secondo, terzo, quarto, quinto e sesto parto, rispettivamente. Infine, per la valutazione della lattazione sono stati presi in considerazione solo gli animali che presentassero almeno 5 TD. Per ciascuna variabile considerata è stato calcolato il coefficiente di variazione fenotipica (CV), come rapporto tra la deviazione standard e la media di ciascun parametro considerato utilizzando il sofware statistico SAS versione 9.4 (SAS Institute Inc., Cary, NC, USA). Il medesimo software è stato utilizzato anche per determinare le correlazioni di Pearson tra i caratteri.
Al fine di modellare la curva di lattazione, sono state create 11 classi da 30 giorni ciascuna, eccetto per la prima (5-30), la penultima (271-310) e l’ultima (311-400), che hanno presentato una numerosità differente. Tenendo in considerazione la stagionalità della specie, il mese di parto è stato incluso come effetto fisso (12 livelli). L’analisi della varianza è stata effettuata secondo il seguente modello misto:
yijklm = μ + Pari + Lacj + Monthk + (Par × Lac)ij + (Par × Month)ik + (Lac × Month)jk + IDl + HTDm + eijklm,
ove:
y è la variabile dipendente (MY, ECM, FC, PC, LC e SCS);
μ è l’intecetta;
Par è l’effetto fisso dell’ordine di parto;
Lac è l’effetto fisso dello jth stadio di lattazione;
Month è l’effetto fisso del kth mese di parto;
(Par × Lac) è l’effetto fisso dell’interazione tra ordine di parto e stadio di lattazione;
(Par × Month) è l’effetto fisso dell’interazione tra ordine di parto e mese di parto;
(Lac × Month) è l’effetto fisso dell’interazione tra stadio di lattazione e mese di parto;
ID è l’effetto random del lth bufalo;
HTD è l’effetto random del mth gruppo;
e è l’errore residuale.
Al termine del processo di editing sono quindi stati analizzati 980.330 TD da 162.776 lattazioni di 72.294 bufale in 341 aziende. I TD analizzati hanno presentato una distribuzione omogenea tra gli anni in merito all’ordine di parto, secondo le seguenti percentuali: 1a lattazione: 28,04%; 2a lattazione: 24,70%; 3a lattazione: 18,53%; 4a lattazione: 13,33%; 5a lattazione: 9,41%; 6a lattazione: 5,99%. Per ciascun anno di campionamento è stata calcolata la media delle caratteristiche quali-quantitative del latte (Tabella 1).

Primo riscontro interessante è stato che le medie produttive registrate nel corso delle analisi del progetto STRABUF sono risultate maggiori dei valori ufficiali medi riportati nel Libro Genealogico. Infatti, le medie ufficiali sono calcolate sulla lattazione standard di 270 giorni ed includono anche aziende al di fuori della Regione Campania. Se vengono presi in considerazione soltanto i dati delle aziende localizzate in Regione Campania, la media ufficiale di MY è di 8,60 kg/day, rispecchiando e valorizzando i risultati ottenuti nella nostra ricerca. Va inoltre sottolineato che sia il trend della produzione di latte (MY) che di ECM sono risultati positivi dal 2013 al 2017: ciò sta a sottolineare un miglioramento fenotipico della popolazione ed anche un miglioramento della redditività per capo. Complessivamente, FC e PC hanno fatto registrare una media dell’8,10%±1,65% e 4,70%±0,44%, rispettivamente, con una variabilità che andava dall’8,00% all’8,26% (FC) e dal 4,79% al 4,74% (PC; Tabella 1). Complessivamente, sia FC che PC hanno mostrato una tendenza non lineare nel corso degli anni e le medie sono risultate sovrapponibili alle statistiche nazionali disponibili (2017; FC = 7,89%; PC = 4,61%).
Dopo il grasso, il lattosio è il solido di latte più abbondante nel latte di bufala, con una concentrazione variabile dal 4,50% al 5,20%. Nelle analisi effettuate in questa sede, LC ha avuto una media del 4,76% ± 0,32%, con un minimo (4,72% ± 0,30%) nel 2017 e un massimo (4,79% ± 0,31%) nel 2013 (Tabella 1). Questo valore è risultato maggiore alla media (4,60%) riportata in uno studio su 650 bufale di razza Mediterranea Italiana [Manuelian et al., 2017], ma simile a quello (4,78%) riportato per altre aziende agricole dell’Italia centrale [Tripaldi et al., 2010]. Il valore medio di SCS è stato più alto nel 2017 e più basso nel 2013 (Tabella 1): in particolare, il valore di SCC medio dell’intero periodo è stato di 223.034 cellule/mL, cioè entro l’intervallo riportato in letteratura per il latte di bufalo [Sharif et al., 2007]. È significativo osservare nelle nostre analisi che circa il 26% dei TD (di 59.008 bufali in 105.233 lattazioni) ha presentato valori di SCC > 200.000 cellule/mL, proposto come soglia per l’identificazione dell’infiammazione della mammella nel bufalo [Moroni et al., 2006].
Per quanto concerne il CV, i risultati sono stati leggermente inferiori rispetto a quelli riportati in letteratura in un dataset più piccolo di bufale di razza Mediterranea Italiana [Manuelian et al., 2017]. Durante i 5 anni, il CV di MY, ECM, FC, PC e LC (Tabella 1) erano leggermente inferiori a quelli riportati per un set di dati più piccolo di bufali italiani [Manuelian et al., 2017]. Come nei bovini, FC ha presentato una variabilità maggiore di PC e LC: ciò è stato confermato dai modelli di lattazione presentati in 2 studi effettuati nella specie bufalina [Moroni et al., 2006; Niero et al., 2018]. Tuttavia, sia PC che LC hanno mostrato una maggiore variabilità rispetto al latte bovino. Infine, come in altre specie lattiero-casearie, SCS ha mostrato il CV più elevato tra tutti i tratti studiati in tutti gli anni di campionamento, con CV minimo e massimo nel 2017 e nel 2013, rispettivamente (Tabella 1).
Il coefficiente di correlazione di Pearson (rp) di tutti i tratti investigati è stato calcolato separatamente per le bufale primipare e per le pluripare (Tabella 2).

In particolare, il valore di SCS è stato sempre correlato negativamente sia con MY che con ECM, con rP tra SCS e MY pari a -0,116 e -0,138 in primipare e pluripare, rispettivamente (Tabella 2). Sulla base dell’approccio di confronto delle correlazioni di Weaver et al., 2013, i due coefficienti di correlazione sono risultati significativamente diversi (p < 0,01), suggerendo che l’associazione tra MY e SCS è più forte nelle pluripare che nelle primipare. Nel complesso, ciò ha confermato che la perdita di latte, a causa di un elevato valore di SCS e della cattiva salute della mammella, tende ad aumentare con l’ordine di parto, così come riportato sia nel bufalo [Aspilcueta-Borquis et al., 2010; Fox et al., 2015] che nel bovino [Costa et al., 2019a; ,Costa et al., 2019b]. A causa del cosiddetto effetto diluizione, la riduzione di MY in corrispondenza di valori di SCS elevati, può essere responsabile della correlazione positiva ma debole di SCS con FC e PC (Tabella 2), tra l’altro descritta in precedenza per la stessa specie [Aspilcueta-Borquis et al., 2010]. Correlazioni negative sono invece state riscontrate sia per le primipare che per le pluripare del MY con FC e PC (Tabella 2), confermando nuovamente che una diminuzione della quantità di latte può portare ad un incremento della concentrazione di grasso e proteine, per un progressivo aumento dei valori di SCS [Cerón-Muñoz et al., 2002].
Al contrario, il contenuto in lattosio (LC) è risultato correlato negativamente con SCS, con coefficiente di correlazione di Pearson pari a -0,28 e -0,30 in primipare e pluripare, rispettivamente (Tabella 2). Il coefficiente negativo tra LC e SCS nelle specie da latte sembra essere dovuto alla maggiore permeabilità dell’epitelio e alle giunzioni più deboli nelle strutture alveolari durante fenomeni infiammatori a carico della ghiandola mammaria: pertanto, la perdita di lattosio (nel sangue e nelle urine) nelle ghiandole mammarie infiammate è responsabile del LC inferiore nel lume alveolare e, di conseguenza, nel latte [Fox et al., 2015; Costa et al., 2019a]. A differenza della concentrazione di grasso e proteine (FC e PC), il contenuto in lattosio (LC) non è influenzato dalla diluizione del latte durante la lattazione ma presenta un andamento opposto legato al ruolo osmotico svolto da questo zucchero nel latte: è questo il motivo per il quale LC tende a seguire il medesimo andamento del MY, aumentando fino al picco di lattazione e presentando una discesa successivamente [Costa et al., 2019b; Costa et al., 2019c; Miglior et al., 2006; Haile-Mariam et al., 2017]. La correlazione tra FC e PC è stata positiva e moderata sia nelle primipare che nelle pluripare (Tabella 2), simile anche a quelle riportate nel bovino da latte (0,36 e 0,48). Infine, come previsto, una forte correlazione è emersa tra produzione di latte (MY) e latte corretto (ECM), che hanno fondamentalmente lo stesso comportamento in termini di magnitudine con gli altri parametri investigati (Tabella 2).
Per quanto concerne l’analisi della varianza, la varianza spiegata dall’effetto random dei TD è stata del 55,28, 17,01, 80,06, 74,81, 68,08, e 84,36% per MY, ECM, FC, PC, LC, e SCS, rispettivamente: questi valori confermano che sia la stagione di campionamento che l’azienda (ad es. il management aziendale) influenzano la variabilità della quantità e delle caratteristiche del latte bufalino. Tutti gli effetti fissi sono altamente significativi per spiegare la variabilità dei tratti studiati. Riguardo all’effetto del mese di parto, è importante ricordare che la specie bufalina è a fotoperiodo negativo, incrementando l’attività riproduttiva con la diminuzione delle ore di luce giornaliera [Campanile et al., 2016; D’occhio et al., 2020]. La stagione, quindi, influenza direttamente la fertilità, come testimoniato dalla distribuzione e frequenza dei parti nel corso degli anni (Figure 1 e 2).
Figura 1. Distribuzione dei parti nel corso dell’anno in bufale primipare (■) e pluripare (▲).
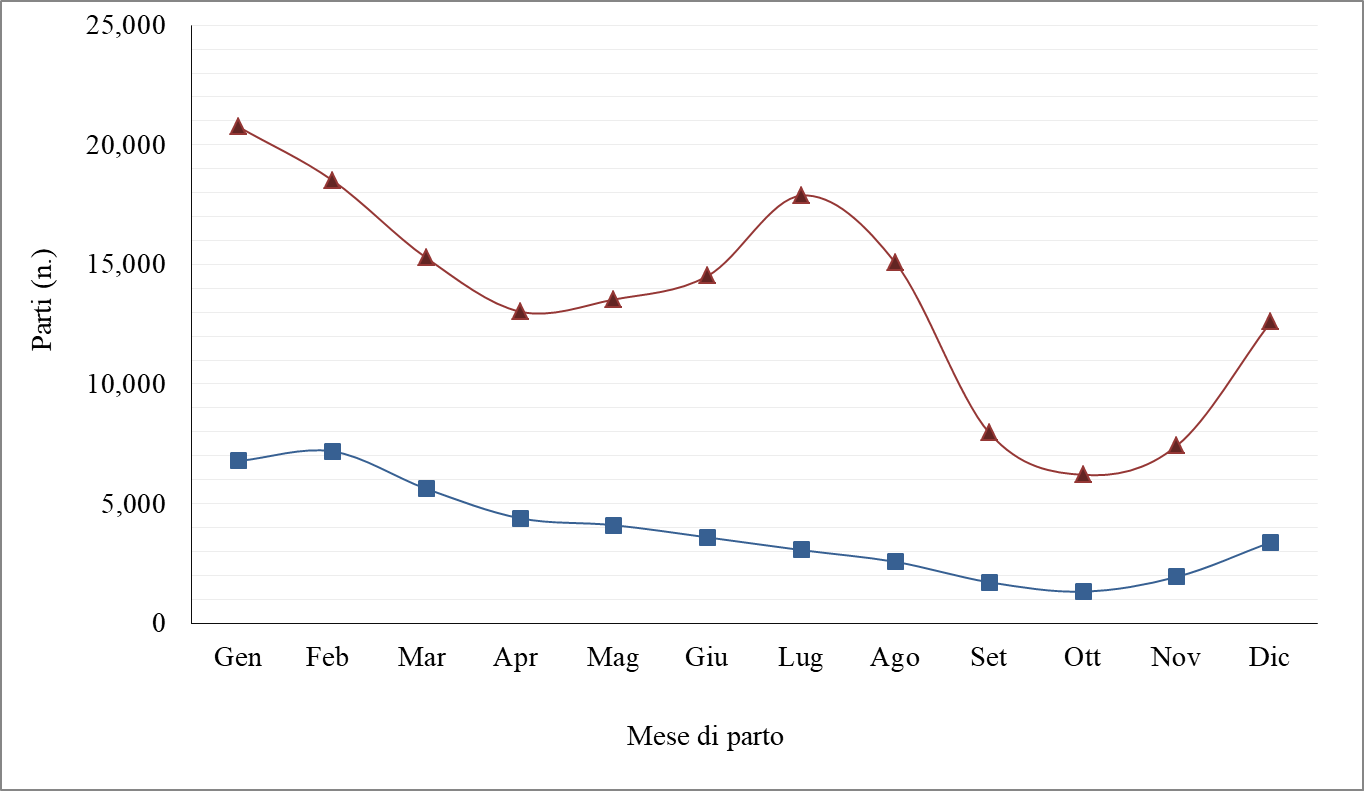
Figura 2. Frequenza dei parti nel corso dell’anno in bufale primipare (■) e pluripare (▲).

La frequenza di parti di bufale primipare è stata massima nel mese di Febbraio ed è diminuita fino ad Ottobre, rispecchiando la tipica caratteristica delle manze bufaline che risultano meno sensibili alla stagionalità riproduttiva rispetto alle pluripare. Basti pensare che il 39% dei parti di Febbraio ed il 37% dei parti di Marzo sono rappresentati da primipare (Figura 3). Al contrario, le bufale pluripare hanno presentato il picco dei parti nel mese di Luglio (>12% del totale dei parti di pluripare). In particolare, le bufale di secondo e terzo parto hanno rappresentato più del 50% dei parti che si sono verificati nei mesi di Luglio e Agosto. Nello stesso periodo, il contributo delle primipare alla totalità dei parti è stato il più basso rispetto al resto dell’anno (Figura 3). È da sottolineare che il trend registrato nelle pluripare non segue la naturale stagionalità della specie [D’Occhio et al., 2020], per l’applicazione di specifiche strategie riproduttive adottate dagli allevatori, quale la tecnica di destagionalizzazione dei parti [D’Onofrio et al., 2019]. Come ampiamente accennato, la bufala è una specie poliestrale stagionale a fotoperiodo negativo. La stagionalità riproduttiva coincide con l’esigenza della specie di dare alla luce la prole in condizioni ambientali favorevoli e con presenza di cibo in abbondanza. La popolazione bufalina è diffusa nelle aree subtropicali-temperate, tra il 31° parallelo Nord ed il 2° parallelo Sud. Come accennato in precedenza, si tratta di una specie tendenzialmente stagionale a fotoperiodo negativo, per cui l’attività riproduttiva in Italia è concentrata nel periodo autunno-vernino (Campanile et al., 2016). Probabilmente questo comportamento è legato a motivi ancestrali, in quanto la culla di origine della specie è la Valle dell’Indo, area nord-equatoriale situata tra India e Pakistan (Zicarelli, 1997) in cui la massima disponibilità di foraggio si ha dopo il periodo delle piogge, che coincide con l’intervallo che va da Luglio a Dicembre-Gennaio. Ovviamente, gli individui nati nelle suddette condizioni, essendo quelli che sopravvivono e che quindi prevalgono numericamente nella popolazione, hanno trasmesso alle generazioni successive le loro peculiari caratteristiche riproduttive.
Figura 3. Contributo delle primipare al totale dei parti nei diversi mesi.

Tutto ciò di cui abbiamo parlato finora è regolato da un meccanismo endocrino: la sorgente luminosa, a seconda dell’intensità e della durata, andrà a stimolare l’asse ipotalamo-ipofisi-gonadi; questo sistema coinvolge la retina, il nucleo soprachiasmatico (NCS), il ganglio simpatico cervicale superiore (SCSG) e, in ultimo, l’epifisi. Gli studi sull’argomento hanno dimostrato che il NCS costituisce un orologio biologico interno che regola il ritmo endogeno circadiano. Gli stimoli che ivi vengono elaborati sono trasmessi attraverso il SCSG all’epifisi, che funziona come trasduttore convertendo le informazioni nervose che derivano dall’alternarsi del ciclo luce/buio. Nella ghiandola pineale il messaggio nervoso stimola il ritmo di secrezione della melatonina che regola attraverso complessi meccanismi l’attività ipotalamo-ipofisaria, e quindi la funzionalità delle gonadi. La luce regola la secrezione di melatonina in due modi: attraverso la stimolazione di un pacemaker circadiano localizzato nel nucleo soprachiasmatico (SCN), che a sua volta controlla l’attività della ghiandola pineale, ed attraverso l’inibizione della produzione di melatonina. La durata della secrezione della melatonina rappresenta quindi un indice endocrino della lunghezza della notte e del giorno (Lincoln et al., 2005). Da alcuni studi, è emerso che anche nella bufala la melatonina rappresenta il segnale endocrino dell’alternanza luce-buio (Zicarelli, 1997). Durante le ore notturne si riscontrano elevate concentrazioni a livello plasmatico di questo ormone, così come in inverno e in autunno rispetto ad estate e primavera.
La stagionalità riproduttiva della specie rappresenta una caratteristica fisiologica che comporta notevoli implicazioni di ordine economico. In Italia, la motivazione economica dell’allevamento bufalino è data dalla produzione della mozzarella, ottenuta esclusivamente con latte di bufala. Tale produzione necessita di una continua disponibilità di latte, o meglio ancora, di una sua maggiore quantità nel periodo primaverile – estivo, quando incrementano i consumi e, al contempo, si verifica una notevole flessione del calendario delle nascite in azienda. È chiaro quindi che la richiesta di latte per far fronte alle esigenze di mercato avviene in contrapposizione alla concentrazione dei parti della specie (Figura 4). Anni addietro si sopperiva alla carenza di latte nel periodo primaverile-estivo utilizzando latte congelato durante il periodo invernale, ma il prodotto finale presentava caratteristiche qualitative ed organolettiche decisamente inferiori rispetto a quello ottenuto da latte fresco. Ciò ha determinato una rilevante flessione del mercato della mozzarella di bufala Campana, che ha indotto i tecnici operanti nel settore a ricercare delle strategie per invertire il calendario dei parti. Da questo nasce l’esigenza di modificare il calendario dei parti intervenendo sull’epoca di accoppiamento mediante la cosiddetta destagionalizzazione dei parti. La tecnica consiste nell’interrompere la promiscuità sessuale nel periodo di massima attività riproduttiva e lasciare il toro nella mandria nel periodo compreso tra Marzo e fine Settembre, in modo da ottenere parti tra la fine di gennaio e inizio agosto (Zicarelli, 1997).
Figura 4. Relazione tra percentuale di parti, produzione di latte e richieste di mercato per la produzione di mozzarella di bufala Campana DOP nel corso dell’anno in condizioni di monta naturale.

E’ indubbio che l’applicazione di tale tecnica comporti una riduzione della fertilità, impedendo gli accoppiamenti degli animali nei periodi in cui la specie si presenta più fertile (cioè quando diminuiscono le ore di luce giornaliera) e inducendoli quando aumentano le ore di luce giornaliera. Proprio per tale motivo sono state elaborate due diverse tecniche di destagionalizzazione: quella graduale e quella drastica. La scelta dell’una o dell’altra tecnica è strettamente dipendente da vari fattori:
- Grado di stagionalità della mandria. Ovvero la percentuale di animali che partoriscono durante il periodo autunno-vernino: se il grado di stagionalità è superiore al 70% si preferisce impiegare una tecnica “soft” che si attuerà nel corso di tre anni, mentre, se il grado è del 40-50%, la tecnica applicata sarà drastica. Una destagionalizzazione graduale in 3 anni, che generalmente si attua in aziende in cui non è stata mai effettuata destagionalizzazione, prevede l’allontanamento dei tori dalla mandria nel primo anno tra fine Ottobre e fine Gennaio, nel secondo tra metà Ottobre e metà Febbraio e nel terzo tra inizio Ottobre e inizio Marzo, al fine di diluire la perdita di fertilità che dovrebbe verificarsi in 1 anno in 3 anni.
- Disponibilità di manze puberi. Come accennato in precedenza, gli animali più giovani risentono meno del fotoperiodo e, pertanto, possono essere più facilmente sottoposti alla tecnica di destagionalizzazione dei parti. Una buona disponibilità di manze, evento non difficilissimo vista la notevole longevità della specie che permette di ridurre il tasso di rimonta, consente di colmare parzialmente la perdita di fertilità che si verifica nelle bufale pluripare per l’applicazione della tecnica. Va in ogni caso sottolineato che le manze possono essere messe in produzione in qualsiasi periodo dell’anno, purchè abbiano raggiunto un peso che oscilli tra i 350-400 Kg di PV.
- Condizioni commerciali della zona. La richiesta di mercato rappresenta il principale fattore da tenere in considerazione prima di applicare la tecnica di destagionalizzazione dei parti. Un caseificio localizzato in una grande città avrà paradossalmente meno richiesta di mozzarella (e quindi di latte) nel mese di Agosto e maggiore a Settembre per il ritorno dalle ferie.
Le aziende che applicano da più anni (più di dieci) la destagionalizzazione, eliminando i soggetti più sensibili al fotoperiodo, presentano un intervallo interparto inferiore rispetto alle aziende che la applicano da meno tempo. La riduzione della fertilità, che può variare dal 10 al 35%, è dovuta alle due principali problematiche riproduttive che si riscontrano nella specie bufalina in Italia: la mortalità embrionale, cioè l’interruzione della gestazione in stadi precocissimi, e l’anaestro, cui vanno incontro nei mesi primaverili soprattutto le bufale pluripare anziane (Zicarelli, 1997). Alcuni studi hanno messo in evidenza come la tecnica determini un aumento dell’interparto, che dipende dall’efficienza riproduttiva della mandria e che è tanto maggiore quanto sono maggiori i problemi concernenti la sfera riproduttiva delle bufale. Da un punto di vista pratico, le manze sono di solito inseminate tra Marzo ed Agosto, mentre le pluripare tra Febbraio e Ottobre. Il maggior prezzo di mercato (generalmente +30% rispetto al resto dell’anno) per il latte nel periodo primaverile-estivo [Presicce, 2007], ripaga delle perdite di fertilità verificatesi per l’applicazione della tecnica [Campanile et al., 2016].
Per quanto concerne l’effetto del mese di parto, è stata registrata solo una lieve variabilità per la maggior parte delle caratteristiche quali-quantitative del latte considerate. La massima produzione sia di latte che di ECM è stata osservata nelle bufale partorite a Marzo, sebbene non siano state rilevate differenze per quelle partorite a Febbraio ed Aprile. Al contrario, le bufale partorite a Luglio, Agosto e Settembre hanno mostrato una produzione significativamente (p<0,05) inferiore. Alcuni studi effettuati in Brasile, al contrario, non hanno messo in evidenza alcuna differenza per quanto concerne la produzione di latte e la composizione durante la stagione delle piogge (Settembre-Febbraio) e la stagione secca (Marzo-Agosto) [Bartocci et al., 2002], mentre in altre prove la produzione di latte registrata nelle bufale partorite durante l’autunno e l’inverno (5.59 ± 0.10 kg/d) è stata significativamente maggiore di quella registrata nelle bufale partorite in primavera ed estate (5.29 ± 0.06 kg/d) [Zotos et al., 2014].
Considerando le caratteristiche qualitative, il contenuto in grasso è stato maggiore nelle bufale partorite nei mesi invernali, mentre quello in proteine è iniziato a crescere nei mesi primaverili e ha raggiunto un picco nelle bufale partorite a Luglio (Tabella 3). Sembra quindi che le bufale partorite a Luglio siano caratterizzate dal maggior contenuto in proteine, probabilmente per la minor pressione in questi animali nella seconda metà della lattazione (e quindi durante i mesi invernali). Pochissime variazioni sono invece state osservate per quanto riguarda il lattosio e lo score delle cellule somatiche (SCS), suggerendo che la variazione di tali caratteri sia probabilmente dovuta ad altri fattori, quali ordine di parto e stadio di lattazione (Tabella 3). Una discussione approfondita sui tratti considerati è stata difficile, a causa della scarsità di lavori in letteratura che studiano l’effetto del mese del parto su FC, PC, LC e SCS. In effetti, solo l’effetto del mese del campionamento è stato ampiamente studiato nei bufali con il massimo (8,54%) ed il minore (6,27%) valore di FC nei campioni di latte campionati a Gennaio e Giugno, rispettivamente. Allo stesso modo, alcuni studi hanno riportato un maggiore FC nel latte campionato in inverno.
Infine, l’interazione tra l’effetto dell’ordine di parto e dello stadio di lattazione per tutti i caratteri studiati è rappresentata nelle figure successive (Figure 5 – 10).

Figura 5. Media della produzione di latte (kg/d) per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.

Figura 6. Media della produzione di latte corretto (ECM, kg/d) per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.

Figura 7. Media del contenuto in grasso (%) per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.

Figura 8. Media del contenuto in proteine (%) per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.
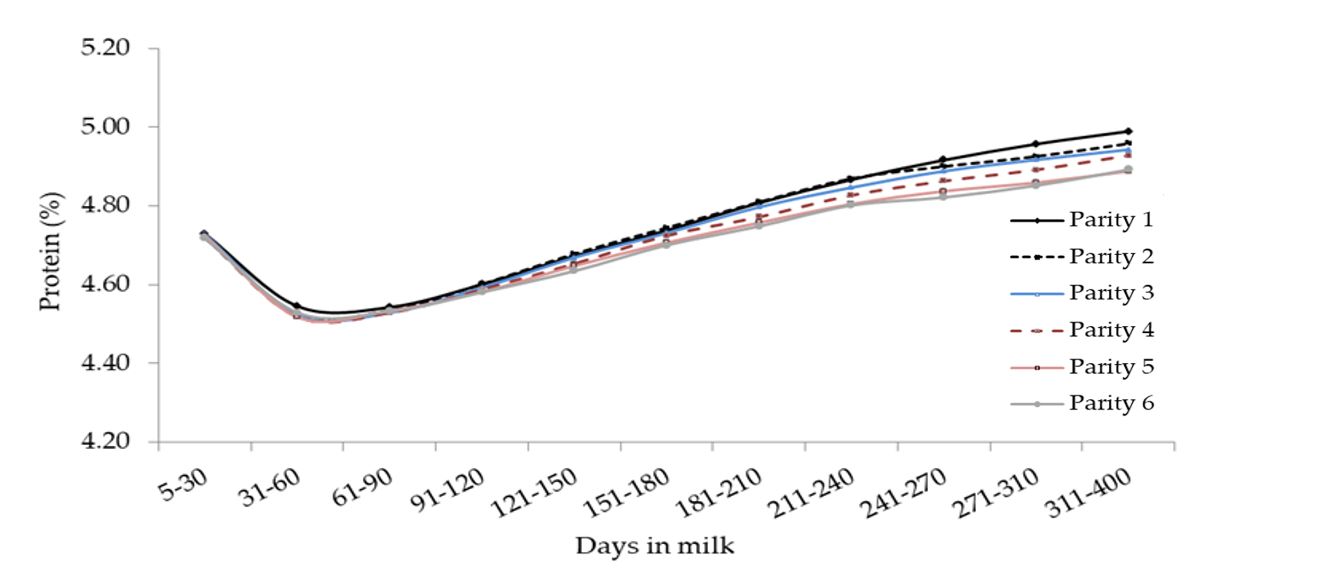
Figura 9. Media del contenuto in lattosio (%) per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.
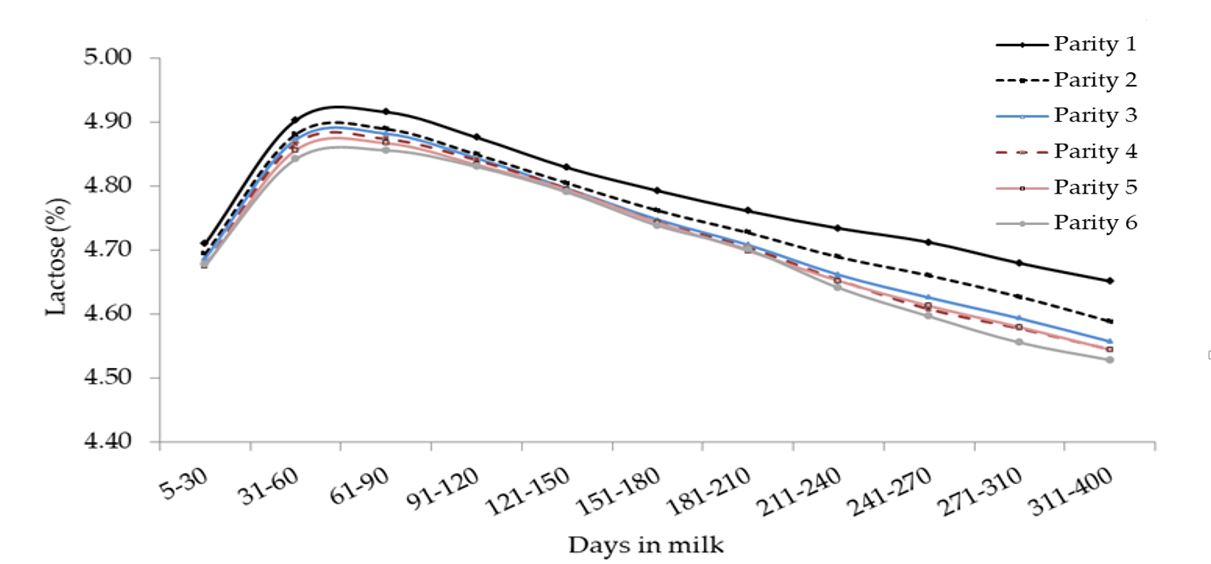
Figura 10. Media di SCS per quanto concerne l’interazione tra ordine di parto e stadio di lattazione.
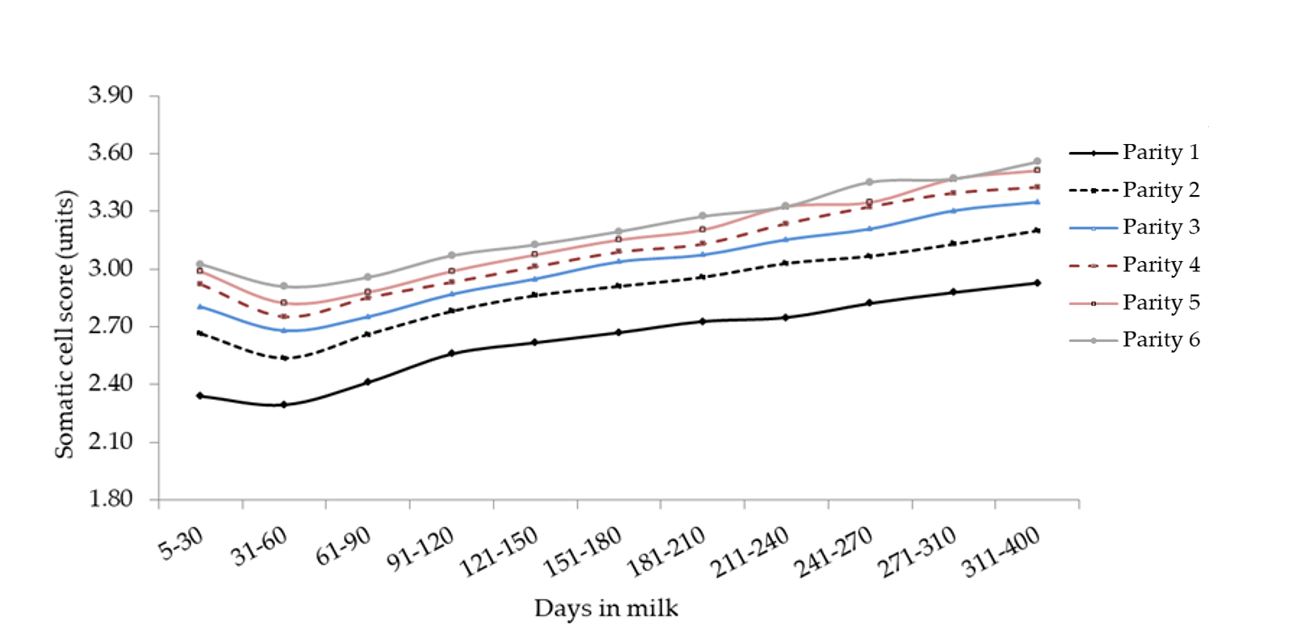
Sia la produzione di latte che quella di ECM sono risultate significativamente differenti nel corso della lattazione tra bufale con diverso ordine di parto, così come osservato in precedenza in bufale Mediterranee [Dürr et al., 2008] e Murrah brasiliane [Cerón-Muñoz et al., 2002] ed altri ruminanti. In particolare, la produzione di latte tendeva ad aumentare dall’inizio della vita produttiva e diminuire successivamente nelle bufale anziane: l’aumento è stato registrato fino alla terza lattazione, mentre si è osservata una lieve diminuzione successivamente. Le ragioni di tale diminuzione vanno ricercate negli effetti combinati dell’invecchiamento dei tessuti e la condizione di sanità sub-ottimale della ghiandola mammaria, come suggerito dall’incremento dello score delle cellule somatiche (Figura 10). La persistenza delle curve di lattazione è risultata simile dal secondo al sesto parto e, a conferma di quanto già riportato in letteratura [Costa et al., 2019b], le bufale primipare hanno fatto registrare una persistenza maggiore delle pluripare (Figura 5). Questo dato potrebbe essere parzialmente spiegato con il ritardo volontario del periodo di inseminazione nelle primipare per applicare la tecnica di destagionalizzazione dei parti.
Nel complesso, le curve di lattazione per quanto concerne il contenuto in grasso, proteine e lattosio sono stati simili a quelli riportati in altri studi effettuati su dataset più ridotti [Cerón-Muñoz et al., 2002]. Generalmente la qualità del latte (in termini di solidi totali) registrata nelle primipare è stata superiore a quella delle altre categorie, mentre il contenuto in lattosio ha mostrato una progressiva diminuzione in base all’ordine di parto pur conservando lo stesso trend, così come descritto anche nei bovini [Costa et al., 2019c, Herve et al., 2018].
Autori: Angela Costa e Massimo De Marchi, Università di Padova.
Tratto da “Phenotypic characterization of milk yield and quality traits in a large population of Water buffaloes“, Costa A., Negrini R., De Marchi M., Campanile G., Neglia G. (2020). Animals; 10: 327 (DOI number: 10.3390/ani10020327)
Bibliografia
- Aspilcueta-Borquis, R. R.; Araujo Neto, F. R.; Baldi, F.; Bignardi, A. B.; Albuquerque, L. G.; Tonhati, H. Genetic parameters for buffalo milk yield and milk quality traits using Bayesian inference. J. Dairy Sci. 2010, 93, 2195 – 2201. https://doi.org/10.3168/jds.2009-2621
- Bartocci, S.; Tripaldi, C.; Terramoccia, S. Characteristics of foodstuffs and diets, and the quanti-qualitative milk parameters of Mediterranean buffaloes bred in Italy using the intensive system: An estimate of the nutritional requirements of buffalo herds lactating or dry. Livest. Prod. Sci. 2002, 77, 45 – 48. https://doi.org/10.1016/S0301-6226(02)00022-2
- Campanile, G.; Neglia, G.; D’Occhio, M. J. Embryonic and fetal mortality in river buffalo (Bubalus bubalis). Theriogenology. 2016, 86, 207 – 213. http://dx.doi.org/10.1016/j.theriogenology.2016.04.033
- Cerón-Muñoz, M.; Tonhati, H.; Duarte, J.; Oliveira, J.; Muñoz-Berrocal, M.; Jurado-Gámez, H. Factors affecting somatic cell counts and their relations with milk and milk constituent yield in buffaloes. Dairy Sci. 2002, 85, 2885 – 2889. https://doi.org/10.3168/jds.S0022-0302(02)74376-2
- Costa, A.; Egger-Danner, C.; Mészáros, G.; Fuerst, C.; Penasa, M.; Sölkner, J.; Fuerst-Waltl, B. Genetic associations of lactose and its ratios to other milk solids with health traits in Austrian Fleckvieh cows. J. Dairy Sci. 2019a, 102, 4238 – 4248. https://doi.org/10.3168/jds.2018-15883
- Costa, A.; Lopez-Villalobos, N.; Sneddon, N. W.; Shalloo, L.; Franzoi, M.; De Marchi, M.; Penasa M. Invited review: Milk lactose – Current status and future challenges in dairy cattle. J. Dairy Sci. 2019c, 102, 5883 – 5898. https://doi.org/10.3168/jds.2018-15955
- Costa, A.; Lopez-Villalobos, N.; Visentin, G.; De Marchi, M.; Cassandro, M.; Penasa, M. Heritability and repeatability of milk lactose and its relationships with traditional milk traits, somatic cell score and freezing point in Holstein cows. Animal. 2019b, 13, 909 – 916. https://doi.org/10.1017/S1751731118002094
- D’Occhio M.J., Ghuman S.S., Neglia G., Della Valle G., Baruselli P.S., Zicarelli L., Visintin J.A., Sarkar M., Campanile G. (2020). Exogenous and endogenous factors in seasonality of reproduction in buffaloes: A review. Theriogenology; 150: 186 – 192. http://dx.doi.org/10.1016/j.theriogenology.2020.01.044
- D’Onofrio N., Balestrieri A., Neglia G., Monaco A., Tatullo M., Casale R., Limone A., Balestrieri M.L., Campanile G. (2019). Antioxidant and anti-inflammatory activities of buffalo milk δ-Valerobetaine. Journal of Agricultural and Food Chemistry; 67 (6): 1702 – 1710. https://doi.org/10.1021/acs.jafc.8b07166
- Dürr, J. W.; Cue, R. I.; Monardes, H. G.; Moro-Méndez, J.; Wade, K. M. Milk losses associated with somatic cell counts per breed, parity and stage of lactation in Canadian dairy cattle. Livest Sci. 2008, 117, 225 – 232. https://doi.org/10.1016/j.livsci.2007.12.004
- Fox, P. F.; Uniacke-Lowe, T.; McSweeney, P. L. H.; O’Mahony, J. A. Dairy Chemistry and Biochemistry. Springer International Publishing, Basel, Switzerland, 2015
- Haile-Mariam, M.; Pryce, J. E. Genetic parameters for lactose and its correlation with other milk production traits and fitness traits in pasture-based production systems. Dairy Sci. 2017, 100, 3754 – 3766. https://doi.org/10.3168/jds.2016-11952 [Corrigendum. J. Dairy Sci. 100, 6008 – 6008; https://doi.org/10.3168/jds.2017-100-7-6008]
- Herve, L.; Lollivier, V.; Quesnel, H.; Boutinaud, M. Oxytocin induces mammary epithelium disruption and could stimulate epithelial cell exfoliation. Mammary Gland Biol. Neoplasia. 2018, 23, 139 – 147. https://doi.org/10.1007/s10911-018-9400-8
- Manuelian, C. L.; Visentin, G.; Boselli, C.; Giangolini, G.; Cassandro, M.; De Marchi, M. Prediction of milk coagulation and acidity traits in Mediterranean buffalo milk using Fourier-transform mid-infrared spectroscopy. Dairy Sci. 2017, 100, 7083 – 7087. https://doi.org/10.3168/jds.2017-12707
- Miglior, F.; Sewalem, A.; Jamrozik, J.; Lefebvre, D. M.; Moore; R. K. Analysis of milk urea nitrogen and lactose and their effect on longevity in Canadian dairy cattle. J. Dairy Sci. 2006, 89, 4886 – 4894. https://doi.org/10.3168/jds.S0022-0302(06)72537-1
- Moroni, P.; Sgoifo Rossi, C.; Pisoni, G.; Bronzo, V.; Castiglioni, B.; Boettcher, P. J. Relationships between somatic cell count and intramammary infection in buffaloes. Dairy Sci. 2006, 89, 998 – 1003. https://doi.org/10.3168/jds.S0022-0302(06)72165-8
- Niero, G.; Currò, S.; Costa, A.; Penasa, M.; Cassandro, M.; Boselli, C.; Giangolini, G.; De Marchi, M. Short communication: Phenotypic characterization of total antioxidant activity of buffalo, goat, and sheep milk. Dairy Sci. 2018, 101, 4864 – 4868. https://doi.org/10.3168/jds.2017-13792
- Presicce, G. A. Reproduction in Water Buffalo. Dom. Anim. 2007, 42 (Suppl. 2), 24 – 32. https://doi.org/10.1111/j.1439-0531.2007.00907.x
- Sharif, A.; Ahmad, T.; Bilal, M. Q.; Yousaf, A.; Muhammad, G. Effect of severity of sub-clinical mastitis on somatic cell count and lactose contents of buffalo milk. Pakistan Vet J. 2007, 27, 142 – 144. http://www.pvj.com.pk/pdf-files/27_3/page%20142-144.pdf.
- Tripaldi, C.; Palocci, G.; Miarelli, M.; Catta, M., Orlandini, S.; Amatiste, S.; Di Bernardini, R., Catillo, G.. Effects of mastitis on buffalo milk quality. Asian-Aust. J. Anim. Sci. 2010, 23, 1319 – 1324. https://doi.org/10.5713/ajas.2010.90618
- Weaver, B.; Wuensch, K. L. SPSS and SAS programs for comparing Pearson correlations and OLS regression coefficients. Behav. Res. 2013, 45, 880 – 895. https://doi.org/10.3758/s13428-012-0289-7
- Zicarelli, L. Reproductive seasonality in buffalo. Third Course on Biotechnologies of reproduction in buffalo. Proc. V° World Buffalo Congress, Caserta, Italia, 13-18 Ottobre 1997: pag. 29-52.
- Zotos, A.; Bampidis, V. Short Communication: Milk fat quality of Greek buffalo (Bubalus bubalis). J. Food Compos. Anal. 2014, 33, 181 – 186. https://doi.org/10.1016/j.jfca.2013.12.004




